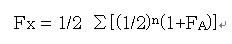 ここで、FXは個体Xの近交係数、nは個体xの父から共通祖先Aを経由して個体xの母までの世代数、FAは共通祖先Aの近交係数を表しています。
ここで、FXは個体Xの近交係数、nは個体xの父から共通祖先Aを経由して個体xの母までの世代数、FAは共通祖先Aの近交係数を表しています。
ある形質について優れた者は優れた者どうし、劣る者は劣る者どうし任意することを同類交配と呼ぶ。 家畜では任意交配されることはなく、同類交配などの人為的な交配が行われる。
また、血縁の近い者どうしを交配することを近親交配と呼ぶ。 家畜においては選抜育種が進むと近親交配に陥りやすい。 逆に、血縁的に遠い者どうしを交配することを異系交配と呼ぶ。
| 遺伝子型 | 頻度 |
| A1A1 | p12 |
| A1A2 | 2p1p2 |
| A2A2 | p22 |
また、対立遺伝子が3つ以上ある場合にも、このハーディーワインベルクの法則は成立する。 さらに、任意交配集団でハーディーワインベルクの平衡状態にあれば、 2つ以上の遺伝子座について考慮する場合には、 各遺伝子型の頻度はそれぞれの遺伝子座の遺伝子型頻度の積となる。 すなわち、A1の頻度をp1、A2の頻度を p2、B1の頻度をr1、B2の頻度をr2とすると、 遺伝子型A1A1B1B1の頻度は p12r12となり、 遺伝子型A1A2B1B2の頻度は 4p1p2r1r2となる。
ただし、1つの遺伝子座に関しては1世代の任意交配でハーディーワインベルクの平衡に達するが、 複数の遺伝子座について考えると、すぐには平衡状態にならない。 特に、2つの遺伝子座が強く連鎖している場合には、何世代も平衡でない状態が続く。 これを連鎖不平衡と呼ぶ。 逆にいうと、1つの遺伝子座については平衡状態と考えられる集団でも、 多数の遺伝子座についてはほとんどの場合、 平衡状態にあるとは仮定できない。
遺伝様式を仮定することによって、各表現型の期待数を算出できる。 これを実際の観察数と比較することによって、遺伝様式の仮定が 正しいかどうかを判定することができる。 統計学的な判定には通常カイ自乗検定を用いる。
なお、近交係数は以下の式で求められる。
ここで、FXは個体Xの近交係数、nは個体xの父から共通祖先Aを経由して個体xの母までの世代数、FAは共通祖先Aの近交係数を表しています。
Σは共通祖先を経由する経路の数だけ足し合わせることを意味しています。
遡って調べる世代数を多くすると、経路の数が増加し、計算量もかかりますが、 現在よりも何世代も遡っても近交係数へ効果は小さいので、血統情報があっても近交係数の計算には、3-5世代までの情報を利用することが一般的です。
FAをゼロと見なした式が書かれている教科書もありますが、黒毛和種のような近親交配が進んだ集団でFAをゼロと見なすのは危険です。
なお、血縁係数は以下の式で求められる。
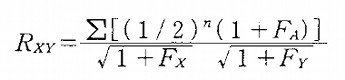 ここで、RXYは個体Xと個体Yの間の血縁係数、nは個体Xから共通祖先Aを経由して個体Yまでの世代数、FAは共通祖先Aの近交係数、
FXは個体Xの近交係数、FYは個体Yの近交係数を示しています。
ここで、RXYは個体Xと個体Yの間の血縁係数、nは個体Xから共通祖先Aを経由して個体Yまでの世代数、FAは共通祖先Aの近交係数、
FXは個体Xの近交係数、FYは個体Yの近交係数を示しています。
Σは共通祖先を経由する経路の数だけ足し合わせることを意味しています。
後述するBLUP法で血縁情報を利用する時には相加的血縁係数行列を用いる。
雄雌の数が異なる場合、集団の有効な大きさNEは NE = 4NmNf / (Nm+Nf) となる。
ここで、Nmは雄の数、Nfは雌の数である。
佐々木義之 著、「動物の遺伝と育種」、朝倉書店

最終更新年月日 2009年7月21日